|
Paneles de Discussión
Paneais de Discussio Comunicaciones libres
Comunicaçoes livres |
Testing protein permeability of dialysis membranes using SDS-PAGEH. MANN, H. MELZERInstitute of Experimental Nephrology. Aachen. Germanyhelmut.mann@post.rwth-aachen.deABSTRACT:
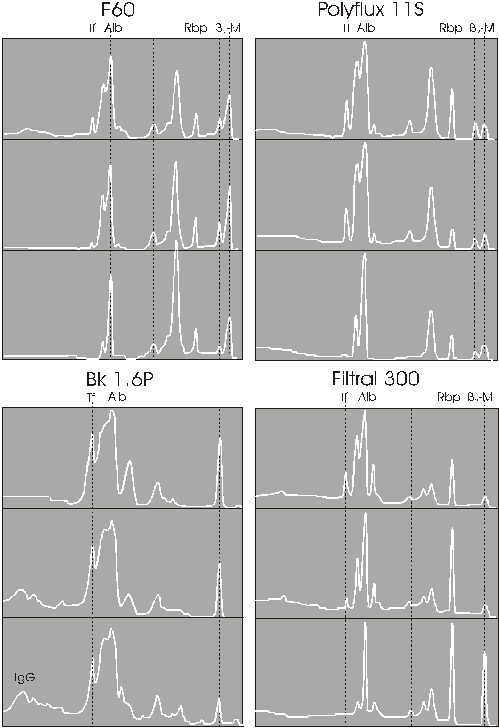
Background: Permeability of dialysis membranes for high molecular weight compounds should be similar to that of the glomerular membrane in order to remove uremic toxins like the human kidney. In order to evaluate permeability of high-flux dialysis membranes SDS-PAGE is applied for examination of filtrate of dialysers during routine dialysis with different membranes. Method: SDS-PAGE analysis is performed with silver staining and consecutive laser densitometry according to the method of Melzer. Results: The protein pattern of filtrate from dialysis membranes is similar to that of the glomerular membrane containing IgG, transferrin, albumin, alpha-1-microglobulin, retinol-binding protein and beta-2-microglobulin. Comparing different membranes there are considerable differences depending on cut-off, charge and adsorption capacity of the particular membrane. In all membranes tested permeability of proteins decreases during one treatment session. Conclusion: Protein permeability of high-flux dialysis membranes is similar to the glomerular membrane but modified according to pore-size, surface charge, adsorption and time on dialysis. In contrast to the glomerular membrane in each of the investigated membranes protein permeability decreases during function. KEY WORDS: Membranes, permeability, hemodialysis, high-flux dialysers SDS-PAGE INTRODUCTION: Uremic toxicity has been detected in non identified molecules of the molecular weight range between 10.000 and 50.000 [1]. These compounds can be found in urine of healthy persons and in serum of patients with uremic syndrome. They are normally excreted by the kidney by filtration through the glomerular membrane. Hence also membranes for artificial kidney therapy should be similar to the glomerular membrane in order to remove these substances from blood. From industry a variety of membranes is provided with a wide range of molecular cut-off. In order to evaluate these membranes with regard to permeability of these molecules, methods are needed for clinical testing during dialysis therapy. Here SDS-PAGE is proposed as a method for testing protein permeability at any time during regular dialysis treatment. METHOD: SDS-PAGE Electrophoretic separation of proteins in filtrate of dialysers is performed by means of ready-to-use gels in the horizontal PhastSystem (Pharmacia, Freiburg, Germany). The continuous polyacrylamid gradient in separating gel is 8-25%, and the degree of cross-netting 2%. The sample used for protein analysis are taken from single filtrate supplemented with sodium azide (final concentration 0.1 g/l). A 10 ml aliquot is centrifuged for 10 min at 3000 rpm. Trichloroacetic (TAC) precipitation is performed in the supernatant, and protein concentrations are determined in the precipitate by the Biuret method. Depending on the protein concentration, electrophoretic protein analysis is performed either in the native filtrate (protein concentration < 0.6 g/l) or in filtrate samples diluted to a protein concentration of 0.6 g/l. Thereafter 0,1 ml of a 5 % SDS buffer solution is added to 0.4 ml filtrate and incubated for 60 min at 37ş C. The final protein concentration of the sample is 0.48 g/l. Then, 1.0 The protein fraction then undergo silver staining in the phast gel according to a modified procedure by Heukeshoven and Dernick [2,4,5,6]. Modified silver staining permits high-resolution separation of the filtrate proteins even at low total protein concentration of < 50 mg/l. Due to the absence of background staining, these stained bands can be transferred into visually recognizable protein patterns by means of laser densitometry (Ultrascan XL Pharmacia, Freiburg, Germany). Proteins are evaluated in a molecular weight range from 10 to 200.000 Dalton. At the same time these patterns can be electronically stored and reproduced at any time, e.g., during long-term observation. The method has been standardized using known concentrations of proteins like albumine, Total protein The first step is the precipitation of the proteins in the filtrate with 20% solution of TCA (trichloracetic acid). The test tubes are stored for at least six hours at 4°C to guarantee precipitation of all proteins. Following storage, the test tubes are centrifuged at 4.0°C for 30 min. at 3000 revolutions/ min. Following decanting of the supernatant, 5.0 ml of Biuret reagent is pipetted onto the protein sediment of each tube, and these were shaken for at least a minute on the rotary mixer until the sediment is completely dissolved in the Biuret reagent. The solutions are measured photometrically after an incubation time of 1 hour. The sample absorbances are measured at 564 nm in the photometer following their transfer in plastic cuvettes. Albumin The filtrate samples were subjected to albumin concentration measurements according to the nephelometry method (DADE BEHRING GmbH/Germany). Filtrate samples and control samples were placed into the nephelometer system for measurement - this is based on the program of albumin concentration in urine (mg/l). ß2-microglobulin (ß2-M) The filtrate samples were subjected to ß2-M concentration measurement according to the Microparticle Enzyme Immunoassay method (MEIA, ABBOTT-Germany). The reaction tubes were placed into the AxSym system for measurement - this is based on the program of ß2-M concentration in urine (mg/l). If the concentration of the sample was below the minimum limitation of measurement, then the sample was subjected to another program which could detect the lower level of ß2-M. Membranes and dialysers The following dialysers have been examined under routine dialysis conditions:  Patients For testing the different dialysers the filtrates are examined in routine dialysis with blood flow of 200 ml/min and dialysate flow of 500 ml/min in unselected patients. Testing change of protein permeability during dialysis therapy the dialysers Filtral 16, F 60, Polyflux S-11 and BK 1,6 F have been investigated in a group of 10 patients alternatively. Samples were taken after 15 min, 30 min and 240 min after start of dialysis therapy. Filtrate sampling Filtrate of the dialysers has been obtained by changing to pure ulrafiltration (Bergström procedure) at a filtration rate of 1000 ml/min. 30 min after changing to pure ultrafiltration a sample from the dialysate compartment of the dialyser is obtained. RESULTS Protein patterns of different membranes In. Fig.1 the different protein patterns in the filtrate of different dialysis membranes are shown. The white line indicates the protein pattern of the particular membrane. The background (black) shows a typical protein pattern of urine of a patient with interstitial kidney disease indicating the proteins which normally permeate the glomerular membrane. For each dialysis membrane also the total protein concentration in the filtrate is indicated. From these membranes the low-flux polysulfone membrane F6 HPS is the most close membrane with no protein in the filtrate. In the cuprophane membrane a very small amount of albumin can be detected by SDS-PAGE probably due to some pin howls in the membrane. The Althane A-18 is considered as a low-flux membrane according to its UF-index. Nevertheless protein in filtrate is relatively high and besides albumin several low molecular weight proteins can be detected in the filtrate. The AN69, Fresenius polysulfone and polyflux S membranes have clear cutt-off at albumin. There is good appearance of ß-2-M and RBP in the filtrate. The new membranes BS, Arylane, FLX-15-GW, APS and Diapes are more open that the former ones and even transferrin appears in the filtrate. The most open membranes are Superflux, SE 18, BK-1,6 F and Altrex-140-G. IN these membranes also IgG can be detected in filtrate. BK 1,6 F and Altrex-140-G have more than 1g/l protein in filtrate. When comparing all these membranes with the protein pattern of interstitial kidney disease it can be stated that there are the same proteins found in the filtrate of these membranes like in ultrafiltrate from the glomerular membrane.
Change of protein pattern during dialysis In four dialysers the change of protein permeability during progression of dialysis therapy has been investigated. As can be seen from Fig.2 in F60, Polyflux and AN69 transferrin which can be detected at the beginning of therapy disappears after 4 hrs of treatment. These membranes remain open for RBP and ß-2-M where in AN69 this peak increases to the end of treatment.. The BK-membrane is the most open membrane which even gets more open to the end of treatment. Because of its high absorption capacity ß-2-M is not detectable.
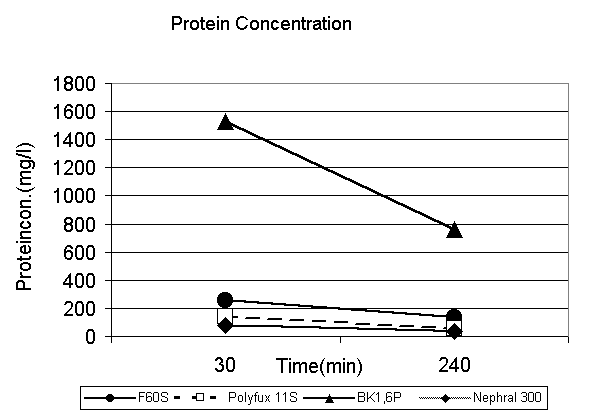
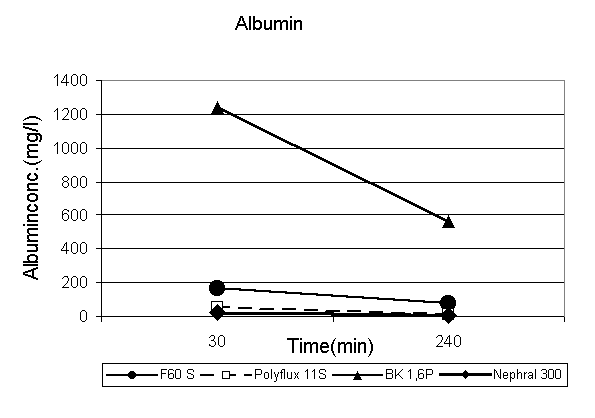
The quantitative changes of protein concentrations in the filtrate during therapy is shown in Fig.3. In each membrane permeability of total protein and albumin decreases during therapy. The higher the initial protein concentration the higher the absolute decrease. In the BK-membrane there is initially no ß-2-M in the filtrate but it increases slightly after 240 min of treatment.
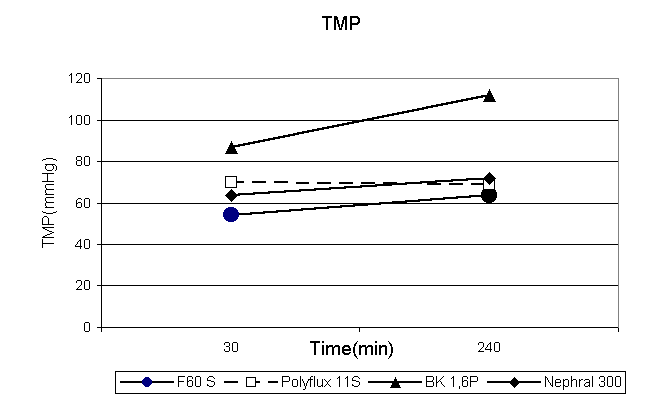
DISCUSSION SDS-PAGE is a method used for detection and separation of proteins according to their molecular weight. In clinical nephrology it is used for diagnosis of glomerular and tubular kidney disease. In all kinds of glomerulonephritis there is increased protein permeability of the glomerular membrane and proteins greater than albumin i.e. immunoglobulins can be found in urine. In all kinds of tubulo-interstitial kidney disease where the tubular cells are unable to reabsorb the normally filtered low molecular weight proteins smaller than albumin like ß-2-microglobulin and retinol binding protein these proteins can be detected in urine indicating filtration of these proteins through the glomerular membrane. Hence the protein pattern in urine of patients with tubulo-interstitial kidney disease indicates protein permeability of the glomerular membrane. In order to evaluate protein permeability of artificial kidney membranes SDS-PAGE has been applied in filtrate of these membranes during routine maintenance dialysis therapy. The results show that filtrate from most high-flux membranes exhibits protein patterns which are very similar to that of the glomerular membrane including albumin, α-1-microglobulin, retinol-binding protein and ί-2-microglobulin. Nevertheless there are considerable differences between different membranes. Whereas low-flux polysulfone is completely impermeable to any protein of more than 10 000 Dalton, some membranes are even open for albumin, transferrin and immunoglobulins. Likewise there are great differences in total protein concentration of different membranes. Some membranes (B1,6 F and Altrex) have protein concentration in filtrate of more than 1g/l which make them not applicable for hemofiltration. On the other hand it can be seen that cellulosic membranes have a broader whereas synthetic membranes have o more clear cut pore size distribution. In cuprophane there is a small amount of albumin detectable at very low total protein concentration and in cellulose diacetate there are even immunoglobulins in the filtrate in contrast to polysulfone, AN69, polyethersulfon where no immunoglobulins are found. Comparing the protein patterns of some of these membranes during progression of dialysis therapy it could be found that in each of the membranes under investigation there is a decrease in total protein content in the filtrate and increase in transmembrane pressure most likely because of adsorption of proteins on the membrane. In most of these membranes transferrin disappears in the filtrate after 4 hrs of treatment. In the PMMA membrane which is known for high adsorptive capacity ß-2-microglobulin is not found in filtrate at the beginning of treatment but later when absorption capacity of the membrane is exhausted it can be seen in the filtrate. Besides total protein also albumin and ß-2-microglobulin concentration in filtrate decreases during therapy in the other membranes. Although sensitivity of SDS-PAGE method as it is performed here for different proteins is not less than other analytical methods ( immuno-assays ) SDS-PAGE gives only relative and not absolute concentrations. So it can not be used for evaluation of quantitative amounts of loss of different proteins. But nevertheless combined with measurement of total protein it gives a good overview of the proteins found in the filtrate and of the molecular cut-off of the different membranes. The results of this investigation show that there is a broad variety of membranes provided from industry for artificial kidney therapy. Most of the so-called high-flux membranes are somehow similar to the glomerular membrane and in principle able to remove so-called middle- molecular weight uremic toxins. The low-flux membranes seem not be suitable for removal of these substances. But there are also membranes whose high protein permeability make them not applicable for hemofiltration. In each of the membranes there is decrease of protein permeability during treatment. The simplicity of the method allows control of protein permeability of dialysis membranes at any time during routine dialysis therapy. SUMMARY SDS-PAGE is a method which can be used for examination of protein permeability of dialysis membranes at any time during dialysis therapy. High-flux membranes exhibit protein patterns in the filtrate which are similar to the glomerular basement membrane. But there are considerable differences between different membranes induced by absorption, molecular cut-off and total protein concentration. Low-flux membranes are unable to remove so-called middle molecular weight uremic toxins. In contrast to the natural kidney in all membranes protein permeability decreases during 4 hours of treatment. 1. Brunner H, Mann H, Essers U, Schultheis R, Byrne T, Heintz R.: Preparative isolation of middle moleculer weight fractions from the hemodialysate of patients with chronic uremia. Artif Organs 1978;2: 375
2. Broklebank T, Cooper EH, Richmond K.: Sodium dodecyl sulphate polyacrylamide gel electophoresis patterns of proteinuria in various renal disease of childhood. Pediatr Nephrol. 1991; 5: 371-375. 3. Weber WH.: Urinary proteins analysis. J Chromatogr. 1988; 429:315-344. 4. Kierdorf H, Melzer H, Mann H, Sieberth HG.: Differentation of proteins in polyacrylamide gels by a modification of silver staining for the phast system and a laserdensitometer. Electrophoresis. 1993; 14: 820-822. 5. Heukeshoven J, Dernick R.: Improved silver staining procedure for fast staining in Phast System development unit. I. staining of sodium dodecyl sulfate gels. Electrophoresis 9: 28-32 (1988) 6. Floege J, Barest A, Schools M, Shaldon S, Koch KM, Smeby LC: Clearance and synthesis rates of ß2-microglobulin in patients undergoing hemodialysis and in normal subjects. J Lab Cline Ed 1991; 18: 153-165 |
 l of the SDS filtrate sample is applied to the phast gel. Electrophoretic protein separation is performed in a cooled separating chamber of the PhastSystem with a current intensity of 10 mA .
l of the SDS filtrate sample is applied to the phast gel. Electrophoretic protein separation is performed in a cooled separating chamber of the PhastSystem with a current intensity of 10 mA . -1-microglobulin (
-1-microglobulin (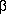 -2 microglobulin (
-2 microglobulin (